

Autor : De Vito, Eduardo L.1,2
1Instituto de Investigaciones MĂ©dicas Alfredo Lanari, Facultad de Medicina, Universidad de Buenos Aires, Buenos Aires, Argentina. 2 Centro del Parque, Cuidados Respiratorios, Buenos Aires, Argentina.
https://doi.org/10.56538/ramr.OKRA7194
Correspondencia : Eduardo Luis De Vito, eldevito@gmail.com
RESUMEN
Todas
las teorĂas sobre los mecanismos de generaciĂłn de disnea tuvieron defensores y
detractores e, interesantemente, con el desarrollo de sofisticadas técnicas
neurofisioÂlĂłgicas y de imágenes funcionales ha sido posible jerarquizar cada
uno de ellos. Todas han sobrevivido al paso del tiempo y ninguna puede explicar
por sĂ sola la disnea en todas las situaciones clĂnicas, lo cual habla de la
naturaleza compleja y multifactorial del fenĂłmeno. El concepto de inadecuaciĂłn
tensión y longitud halló en las últimas décadas un sustento con nuevas
evidencias a su favor. En particular, con el hallazgo de las vĂas involucradas
y con la aplicaciĂłn de conocimientos neurofisiolĂłgicos, la teorĂa de la
inadecuaciĂłn tensiĂłn y longitud se verĂa refinada con la descarga corolaria o copia eferente. Esta descarga corolaria o copia eferente es un atributo básico del
sistema nervioso, que se encuentra en el reino animal, desde los invertebrados
a los primates y en la especie humana. Este artĂculo está dedicado a la historia
de la copia eferente y su incorporaciĂłn como hipĂłtesis para explicar la disnea,
la más aceptada en la actualidad.
Palabras
clave: Disnea,
Mecánica respiratoria, Descarga corolaria, Copia
eferente
ABSTRACT
All the theories about the
mechanisms of generation of dyspnea had defenders and detractors and,
interestingly, with the development of sophisticated neurophysiological
techniques and functional imaging, it has been possible to rank each one of
them. All have survived the passage of time and none can singularly explain
dyspnea in all cliniÂcal situations, showing the complex and multifactorial
nature of the phenomenon. The concept of length-tension inappropriateness has
found support in recent decades with new evidence in its favor. Specially with the discovery of the pathways involved and
with the application of neurophysiological knowledge, the length-tension
inappropriateÂness theory would be refined with the corollary discharge or
efferent copy. This corolÂlary discharge or efferent copy is a basic attribute
of the nervous system found in the animal kingdom, from invertebrates to
primates and in the human species. This article is dedicated to the history of
the efferent copy and its incorporation as a hypothesis to explain dyspnea,
which is currently the most accepted one.
Key words: Dyspnea, Breathing Mechanics, Corollary Discharge, Efferent
Recibido: 26/11/2022
Aceptado: 31/07/2023
INTRODUCCIĂ“N
ÂżPor
quĂ© no podemos hacernos cosquillas a noÂsotros mismos? ÂżPor quĂ© un pez elĂ©ctrico
no se electrocuta? ¿Por qué la tremenda vibración de las patas del grillo no lo
perturba? ¿Por qué los murciélagos no confunden sus sonidos con los de otros?
y, en definitiva, ¿por qué tenemos disnea? Porque existe la copia eferente (CE,
efferent copy),
o descargas corolarias (DC, corollary
discharge).1
Una
CE o DC es un atributo básico del sistema nervioso que se encuentra en el reino
animal, desde los invertebrados a los primates y en la especie humana.1 Este artĂculo
está dedicado a la historia de la CE y su incorporación como hipótesis para
explicar la disnea, la más aceptada en la actualidad.
Cuando
el sistema motor envĂa una señal a un mĂşsculo, tambiĂ©n envĂa una copia interna de
la señal que no sale del sistema nervioso central (SNC). Esta señal interna se
llama “copia eferente” o «descarga corolaria». Esta
CE o DC se compara con la entrada sensorial o referencia (reafferent),
que proviene del mĂşsculo en movimiento. Ahora bien, si la CE/DC y la reaferencia son iguales, significa que el movimiento
programado es igual al movimiento efectuado. Esto evita percepciones autoinducidas innecesarias (Figura 1).
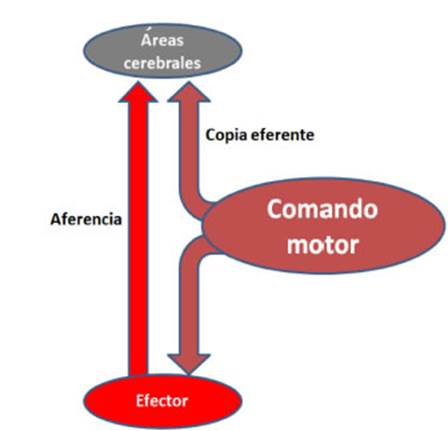
La
orden que se envĂa desde una regiĂłn motora del sistema nervioso central
(comando motor) se copia y se envĂa a otras regiones del SNC antes de que se
produzca el movimiento. Luego, el efector (por ejemplo, mĂşsculo) envĂa
informaciĂłn aferente al SNC, donde ambas señales se comparan. Si amÂbas señales
son iguales (movimiento programado = movimiento efectuado), no hay percepciones
autoinducidas innecesarias.
HISTORIA DE LA COPIA
EFERENTE
El
primero en proponer la existencia de la CE fue Hermann
von Helmholtz a mediados del siglo XIX: el SNC
necesitaba crear una CE desde el comando motor que controla los mĂşsculos
oculares a fin de ayudar al cerebro a determinar la ubicaciĂłn de un objeto con
relaciĂłn a la cabeza. Él acuñó el tĂ©rmino «psicofĂsica» y estableciĂł una
relaciĂłn precisa y no lineal entre la magnitud de los estĂmulos fĂsicos y la
intensidad percibida. Helmholtz allanĂł el camino para
desarrollar las leyes psicofĂsicas de Weber y de Stevens.
El
concepto inicial de CE fue desestimado duÂrante 75 años desde que, en 1920, Sir
Charles Scott Sherrington (Premio Nobel Medicina
1932) criticĂł duramente las ideas de Von Helmholtz.2
Hubo que esperar a mediados del siglo XX cuando Erich von Holst y Horst Mittelstaedt, en
1950, describieron el principio de reaferencia
para explicar cĂłmo un orÂganismo es capaz de separar un estĂmulo sensorial reaferente (autogenerado) de un estĂmulo exaferenÂte (generado externamente).3 Este concepto
aportĂł al entendimiento de los procesos interactivos entre el SNC y su
periferia y fue galardonado con un total de 2973 citaciones.
Fue
Roger Wolcott Sperry
(Premio Nobel de FisiologĂa 1981) tras su investigaciĂłn sobre el reÂflejo optocinĂ©tico del nistagmus que
introdujo el concepto de DC y es considerado el creador de ese tĂ©rmino.4 Su artĂculo
fue citado 1636 veces. La CE ha sido implicada en la falta de disnea en pacienÂtes
con COVID-19,5 una hipĂłtesis
que merece ser explorada con mayor detalle.
Diferencias entre
copia eferente y descarga corolaria
Mediante
diferentes lĂneas experimentales, Von Holst y
Mittelstaedt3 se refirieron
esencialmente al concepto “copia eferente”, mientras que su conÂtemporáneo
Sperry4 acuñó el
concepto “descargas corolarias”. El primer concepto
implica una copia real del comando motor (la eferencia)
que se dirige a los mĂşsculos. Este tĂ©rmino parecĂa apropiado para las preguntas
que Von Holtz y Mittelstaedt
abordaban en invertebrados y para el análisis geÂneral del procesamiento
sensorial que tiene lugar en relaciĂłn con la descarga motora. Sin embargo, se
ha hecho evidente que la conexión entre áreas motoras y sensoriales puede
producirse en varios niveles de control motor.
Mediante
estudios en peces, Sperry4 utilizĂł el
segundo concepto, “descargas corolarias”, para
denotar las señales motoras que se influyen en el procesamiento sensorial, pero
su concepciĂłn era menos especĂfica en cuanto a dĂłnde deberĂa surgir la descarga
motora a las vĂas sensoriales. De manera que los tĂ©rminos tienen diferente
hisÂtoria, y algunas diferencias en cuanto al nivel de complejidad, pero suelen
ser tomados como interÂcambiables. Para los propĂłsitos de este artĂculo
serán mencionados en forma indistinta.
En
las décadas venideras, el concepto de “copia eferente” se ampliará
notablemente. Poulet y cols. sugirieron
el uso de DC como un concepto amplio para abarcar las señales neuronales que se
generan en los centros motores y que no se utilizan directaÂmente para generar
la actividad motora en curso, y a menudo actĂşan para modular el procesamiento
sensorial.6
CLASIFICACIĂ“N
TAXONĂ“MICA DE LAS COPIAS EFERENTES O DESCARGAS COROLARIAS
ÂżCĂłmo
se conecta el procesamiento sensorial en invertebrados y la disnea en humanos?
ÂżQuĂ© tipo taxonĂłmico de copia interna es la que produce disÂnea? Crapse y Sommer sugirieron una
clasificaciĂłn taxonĂłmica funcional de las copias eferentes para todo el reino
animal.1 Las descargas
corolarias pueden clasificarse globalmente en
categorĂas de orden inferior y superior de acuerdo con la funciĂłn y el impacto
operativo de la señal.
La
señalización de orden inferior es ubicua, ya que es necesaria para todo
animal que estĂ© equiÂpado con sistemas sensoriales y motores. En ese ámbito,
las descargas corolarias son un mecanismo
discriminatorio que evita las respuestas desadapÂtativas
y la saturaciĂłn sensorial al restringir o filtrar la informaciĂłn. El grillo no
se aturde (y asà puede escuchar otros ruidos del entorno) y el pez eléctrico no
se electrocuta a sĂ mismo.
Los
monos titĂes, cuando aĂşllan, se encuentran con el mismo problema que los
grillos: en prinÂcipio, los sonidos que emiten deberĂan afectar a su audiciĂłn.
Se puede observar un mecanismo de protecciĂłn en la corteza auditiva primaria
del titĂ, donde se suprimen muchas neuronas durante las autovocalizaciones.
La supresiĂłn comienza unos 200 ms antes de la vocalizaciĂłn y continĂşa durante
toda su duraciĂłn. Este podrĂa ser un caso en el que la DC interconecta áreas
motoras y sensoriales que ocupan ámbitos comparables de una vĂa senÂsoriomotora.1
La
señalización de orden superior participa en dos tipos de funciones. En
el lado perceptual, facilita la interpretaciĂłn contextual de la inforÂmaciĂłn
sensorial (análisis) y la construcción y mantenimiento de una
representaciĂłn interna de esta informaciĂłn (estabilidad). En el lado sensorimotor, facilita la adquisiciĂłn de nuevos
patrones motores (aprendizaje) y la ejecuciĂłn de secuencias de
movimiento rápido (planificaciĂłn). Este tipo de descargas corolarias permite que esÂtructuras cerebrales especĂficas
lleven a cabo los ajustes apropiados en anticipaciĂłn de la entrada
sensorial. Cada murciélago solo escucha su sonido y no el de otros, y les
permite construir una repreÂsentaciĂłn cohesiva del mundo. Hasta el momento,
las DC de orden superior se han identificado solo en vertebrados.
No
hay un solo tipo de DC, sino que existen numerosos subtipos que corresponden
tanto a los niveles anatĂłmicos de la fuente como del objetivo y de las
utilidades funcionales.1 Como se puede
observar, esta taxonomĂa ilustra el importante punto de que, aunque la
concepciĂłn original de DC de Sperry4 concuerda con
el flujo general de informaciĂłn de los sistemas motores a los sistemas
sensoriales en todo el reino animal, parece ser inapropiadamente simple usar un
solo concepto para describir la señal.
IDENTIFICACIĂ“N DE LAS
VĂŤAS DE DESCARGA COROLARIA
Las
neuronas que median en estas señales han sido difĂciles de identificar. La
primera evidencia fue una sola interneurona multisegmental de DC responsable de la inhibición presináptica y postsináptica de
las neuronas auditivas en el canto de los grillos (Gryllus
bimaculatus).6
Unas estructuras similares se hallaron en el renacuajo, el
cangrejo de rĂo y la babosa marina Aplysia.
Los estudios en estas especies hacen a la comprensión clásica de las DC:
proyectan y se dirigen a regiones involucradas en el procesamiento de
informaciĂłn reaferente.
Pero
el procesamiento sensorial es altamente dinámico, que tiene en cuenta el estado
de comporÂtamiento del animal. De manera que el análisis de las vĂas sensoriales
en preparaciones bajo anestesia o en reposo podrĂa no proporcionar una imagen
completa del procesamiento sensorial. Quizás, en términos evolutivos, la DC
primero moduló la actividad real y, luego, en cerebros más complejos, también
se dirigiĂł a las regiones involucradas en la integraciĂłn sensorial-motora o la
planificaciĂłn motriz.7
Nuestros
mĂşsculos son sensibles y esto incluye a los mĂşsculos respiratorios. En otras
palabras, recibimos señales sensoriales de músculos que llegan a la conciencia
y nos dicen lo que sucede en esos músculos de la misma manera que las señales
sensoriales de la piel nos dicen lo que sucede allĂ. Los estudios en
animales demostraron que una copia del impulso motor respiratorio se transmite
al mesencéfalo y al tálamo.1-8
DISNEA Y DESCARGA
COROLARIA
Fue
en 1978 con un artĂculo que mereciĂł más de 1000 citas en el que McClosky y cols. propusieron que las
señales corolarias o CE provenientes de los centros
respiratorios del tronco pueden transmitirse a los centros cerebrales
superiores y dar lugar a un conocimiento consciente del comando del motor de
salida; esto puede jugar un papel importante en la formaciĂłn de la sensaciĂłn de
disnea.11
El
concepto de DC es el más ampliamente aceptado para explicar el origen de la
sensaciĂłn de esfuerzo inspiratorio y la disnea.12-14 El esquema propuesto para el sistema
respiratorio es esenÂcialmente el mismo que el descrito en la Figura 1. A
diferencia de los receptores para el dolor, las aferencias que se proyectan a
los centros cerebrales superiores para cotejar con la CE son diversas.15 Además, el
sistema respiratorio tiene un comando motor automático (troncoencefálico)
y voluntario (corteza motora). Estas DC de diferentes fuentes muy probablemente
dan lugar a diferentes senÂsaciones.15
De manera que, en nuestra opiniĂłn, la disnea no es meramente una
copia en papel carbĂłnico del dolor.
Las
DC en el aparato respiratorio con su doble inervaciĂłn involuntaria y voluntaria
se observan en la Figura 2. Durante la respiraciĂłn involuntaria los centros
respiratorios envĂan una CE a la corteza sensorial mientras que, durante
esfuerzos respiraÂtorios voluntarios, es la corteza motora la que envĂa la
copia. A su vez, los mĂşsculos respiratorios envĂan aferencias a la corteza
sensorial. Cuando existe una correspondencia adecuada entre el comando motor y
la informaciĂłn aferente entrante de los receptores sensoriales, no debe haber
sensaciĂłn de disnea (Figura 2). Por el contrario, cuando no hay coincidencia,
el desacoplamiento neuromecánico resultante contribuye
a la gĂ©nesis de la disnea. Este intercambio entre el comando motor y la corÂteza
sensorial es el mecanismo actualmente más aceptado por el cual se toma
conciencia del esfuerzo de la respiraciĂłn.
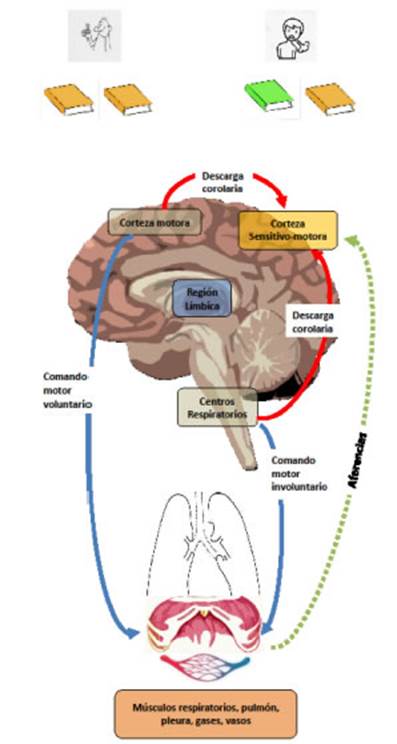
Si
ambas copias (eferente y reaferente) son equiÂvalentes
(igual color), no hay disnea; si las copias difieren (diferente color), la
disnea tiene lugar.
¿Por qué nuestra respiración
no es habitualmente autopercibida?
La
corteza sensorial también recibe aferencias de eventos que se producen en el
tĂłrax y en los mĂşsculos respiratorios y procesa la informaciĂłn.14
Cuando recibe la CE, la corteza sensorial se adapta en consecuencia
para minimizar, eliminar o comÂpensar las consecuencias sensoriales del
movimienÂto. Debido a esta estrategia general, respirar en condiciones normales
es un proceso inconsciente1 (y
dentro de ciertos lĂmites de ventilaciĂłn).
¿Cuál es la relación
entre la inadecuada tensiĂłn-longitud de Campbell y la copia eferente?
En
realidad, una disociación entre el comando motor y la respuesta mecánica del
sistema resÂpiratorio recuerda la teorĂa de la “inadecuaciĂłn de la
longitud-tensión” de Campbell y Howell en la década
de 1960.15 La teorĂa se
ha generalizado para incluir no solo la informaciĂłn que surge en los mĂşsculos
respiratorios, sino la que emana de los receptores a lo largo de todo el
sistema respiratorio y ha recibido diversos términos.15-19
–
Disociación neuromecánica.
–
DisociaciĂłn eferente-reaferente.
–
InadecuaciĂłn tensiĂłn-longitud.
–
DisociaciĂłn neuroventilatoria.
–
Discordancia (mismatch) aferente.
–
Desacople neuromecánico.
–
DisociaciĂłn neuromuscular.
¿Qué condiciones
respiratorias pueden tener disociaciĂłn entre la informaciĂłn eferente y aferente
y producir disnea?
Además
de los hallazgos neurofisiolĂłgicos menÂcionados, diversos datos experimentales
y obserÂvaciones clĂnicas son consistentes con el concepto de inadecuaciĂłn eferente-aferente.20-25
Tanto
en pacientes como en sujetos sanos, la supresiĂłn temporal de la ventilaciĂłn
durante el habla o la comida causa una falta de coincidencia entre el comando
motor respiratorio y el moviÂmiento esperado.
Cuando
los sujetos normales respiran CO2,
su ventilaciĂłn aumenta y la mayorĂa experimenta disnea. Sin embargo, si se
reduce la ventilaciĂłn, pero el CO2 se mantiene
constante, los sujetos inÂforman un aumento marcado en la intensidad de la
falta de aire, a pesar de que el impulso quĂmico para respirar no ha cambiado.
Cuando
los sujetos normales se ven obligados a respirar a un flujo inspiratorio
diferente del que han elegido como el más cómodo, experimentan una sensación de
sed de aire.
Puede
producirse un fenómeno análogo en pacientes que reciben ventilación mecánica y
deÂsadaptaciĂłn al respirador.
Todo
esto sugiere que, bajo un conjunto dado de condiciones, el cerebro «espera» un
cierto patrĂłn de ventilaciĂłn y retroalimentaciĂłn aferente asociada y que las
desviaciones de este patrĂłn causan o intensifican la sensaciĂłn de disnea.
ÂżExiste alguna
relaciĂłn entre la descarga corolaria y falta de
disnea en pacientes con COVID-19?
La
enfermedad por COVID-19 es sorprendente e intrigante en diversos aspectos. Una
de sus caracÂterĂsticas relevantes es que es posible reconocer la falta de
disnea en la mayorĂa de los casos. Si los mecanismos fisiopatolĂłgicos del
desarrollo de la disnea todavĂa no se comprenden bien, no deberĂa sorprendernos
por nuestro conocimiento limitado de los mecanismos de la disnea en la enfermeÂdad
COVID-19.5 En algunas
series, los sujetos intubados y ventilados presentaron taquipnea y taquicardia.26-28 En un estudio
retrospectivo, la disnea y la opresiĂłn torácica fue mucho más coÂmĂşn en
pacientes fallecidos.29 La disnea fue
uno de los predictores asociados de enfermedad grave y muerte.30
Para comprender la ausencia de disnea en COVID-19, el foco
principal está en los fenotipos que muestran hipoxia grave y una distensibilidad del sistema respiratorio casi normal. En el
distrĂ©s respiratorio por neumonĂa por COVID-19, el
sisÂtema respiratorio (distensibilidad transpulmonar y driving
pressure) se informĂł seudonormal.31, 32 Desde la perspectiva de la fisiopatologĂa, que
no excluye el efecto neurotóxico directo del virus y una respuesta sistémica en
el contexto infeccioso, sino más bien lo engloba, la falta de disnea en la
enfermedad por COVID-19 puede explicarse por una adecuaciĂłn en la
corteza sensorial del cerebro de las dos señales procedentes del comando del
motor y la periferia vĂa DC.5
CONCLUSIONES Y
PROYECCIONES TERAPÉUTICAS
Todos
los animales, desde el humilde nematodo hasta el primate cognitivamente avanzado,
requieÂren el tipo de señalizaciĂłn que permite las DC, que protegen de
percepciones autoinducidas innecesaÂrias. TodavĂa
estamos en una etapa embrionaria de investigaciĂłn de las DC en el reino animal.
No obstante, el intercambio entre el comando motor y la corteza sensorial es
el mecanismo actualmente más aceptado por el cual se toma conciencia del
esfuerzo de la respiraciĂłn. Las vĂas neuronales han sido identificadas. En
realidad, una disociación entre el comando motor y la respuesta mecánica del
sistema respiratorio recuerda la teorĂa de la «inadecuaciĂłn de la
longitud-tensiĂłn» de CamÂpbell y Howell en la dĂ©cada
de 1960. El objetivo futuro será descubrir cómo las DC influyen en la
percepciĂłn. Los experimentos hasta ahora han demostrado que la inactivaciĂłn de
las vĂas de DC puede alterar el comportamiento, y los cambios perceptuales
subrepticios pueden justificar estos cambios de comportamiento. Estos
conocimientos pueden ser muy relevante para el alivio
de la disnea refractaria.
PUNTOS CLAVE
•
Datos clĂnicos, experimentales, neurofisiolĂłgicos y las observaciones clĂnicas
avalan el concepto de falta de adecuación tensión-longitud, disociación neuromecánica o disociación eferente reaferente
(CE/DC) como núcleo central en la génesis de la disnea.
•
El concepto central es que, bajo un conjunto dado de condiciones, el cerebro
«espera» un cierto paÂtrĂłn de ventilaciĂłn y retroalimentaciĂłn aferente
asociada; las desviaciones de este patrĂłn causan o intensifican la sensaciĂłn de
disnea.
•
Es necesario profundizar el rol de la DC en la falta de disnea en la enfermedad
por COVID-19 asĂ como en otras condiciones.
BIBLIOGRAFĂŤA
1. Crapse
TB, Sommer MA. Corollary discharge
across the animal kingdom. Nat Rev Neurosci
2008;9:587-600. https://doi.org/10.1038/nrn2457
2. Efference
copy [Internet]. Available from:
https://en.wikipedia.org/w/index.php?title=Efference_copy&oldid=841428446).
3. Von Holst E, Mittelstaedt H. Das Reafferenzprinzip:
Wechselwirkungen zwischen Zentralnervensystem und Peripherie.
Sci Nat 1950;37:464-76.
https://doi.org/10.1007/BF00622503
4. Sperry RW. Neural
basis of the spontaneous optokinetic response
produced by visual inversion. J Comp Physiol Psychol 1950;43:482-9. https://doi.org/10.1037/h0055479
5.
De Vito EL. Possible Role of Corollary Discharge in Lack of
Dyspnea in Patients With COVID-19 Disease [Internet].
Front Physiol 2021;12.
http://dx.doi.org/10.3389/fphys.2021.719166
6. Poulet
JF, Hedwig B. The cellular basis of a corollary disÂcharge.
Science. 2006;311: 518-22.
http://dx.doi.org/10.1126/science.1120847.
7. Poulet
JF, Hedwig B. New insights into corollary discharges mediated by identified
neural pathways. Trends Neurosci 2007;30:14-21. https://doi.org/10.1016/j.tins.2006.11.005
8. Chen Z, Eldridge FL, Wagner
PG. Respiratory-associated rhythmic firing of midbrain neurones
in cats: relation to level of respiratory drive [Internet]. The Journal of
Physiology 1991;437:305-25.
http://dx.doi.org/10.1113/jphysiol.1991.sp018597
9. Chen Z, Eldridge FL, Wagner PG.
Respiratory-associated thalamic activity is related to level of respiratory
drive [Internet]. Respiration Physiology 1992; 90:99-113.
http://dx.doi.org/10.1016/0034-5687(92)90137-l
10. Matthews PBC. Where Does
Sherrington’s “Muscular Sense” Originate? Muscles, Joints, Corollary
Discharges? [Internet]. Annual
Review of Neuroscience 1982; 5: 189-218.
http://dx.doi.org/10.1146/annurev.ne.05.030182.001201
11. McCloskey DI. Kinesthetic sensibility. Physiol
Rev 1978;58:763-820. https://doi.org/10.1152/physrev.1978.58.4.763
12. Spengler CM, Banzett RB, Systrom DM, Shannon
DC, Shea SA. Respiratory sensations during heavy exercise in
subjects without respiratory chemosensitivity.
Respir Physiol 1998;114:65-74. https://doi.org/10.1016/S0034-5687(98)00073-5
13. Booth S, Dudgeon D. Dyspnoea in Advanced Disease: A Guide to Clinical
Management. Oxford University Press; 2006. 271 p.
https://doi.org/10.1093/acprof:oso/9780198530039.001.0001
14. Fukushi
I, Pokorski M, Okada Y. Mechanisms underlying the
sensation of dyspnea. Respir Investig
2021;59:66-80.
https://doi.org/10.1016/j.resinv.2020.10.007
15. Campbell EJ, Howell JB. The sensation of breathlessness. Br Med Bull 1963;19:36-40.
https://doi.org/10.1093/oxfordjournals.bmb.a070002
16. Parshall
MB, Schwartzstein RM, Adams L, et al. An OfÂficial
American Thoracic Society Statement: Update on the Mechanisms, Assessment, and
Management of Dyspnea. Am J Respir Crit Care Med 2012;185:435-52.
https://doi.org/10.1164/rccm.201111-2042ST
17. O’Donnell DE, Webb KA. Exertional
breathlessness in patients with chronic airflow limitation. The role of lung hyperinflation. Am Rev Respir
Dis 1993;148:1351-7.
https://doi.org/10.1164/ajrccm/148.5.1351
18. Banzett
RB, Lansing RW, Brown R, Topulos GP, Yager D, Steele SM, et al. “Air hunger” from increased PCO2 persists
after complete neuromuscular block in humans. Respir Physiol 1990;81:1-17.
https://doi.org/10.1016/0034-5687(90)90065-7
19. Nishino T. Dyspnoea: underlying mechanisms and treatment. Br J Anaesth 2011;106: 463-74. https://doi.org/10.1093/bja/aer040
20. Manning HL, Molinary EJ, Leiter JC. Effect of inspiratory flow rate on respiratory sensation and
pattern of breathing. Am J Respir
Crit Care Med 1995; 151: 751-7.
https://doi.org/10.1164/ajrccm/151.3_Pt_1.751
21. Manning HL, Schwartzstein RM. Pathophysiology of DysÂpnea [Internet]. N
Engl J Med 1995;333:1547-53.
http://dx.doi.org/10.1056/nejm199512073332307
22. Chonan
T, Mulholland MB, Cherniack NS, Altose
MD. EfÂfects of voluntary constraining of thoracic
displacement during hypercapnia. J Appl Physiol 1987;63:1822-8. https://doi.org/10.1152/jappl.1987.63.5.1822
23. Schwartzstein
RM, Simon PM, Weiss JW, Fencl V, WeinÂberger SE.
Breathlessness induced by dissociation beÂtween ventilation and chemical drive.
Am Rev Respir Dis 1989;139:1231-7.
https://doi.org/10.1164/ajrccm/139.5.1231
24. Manning HL, Shea SA, Schwartzstein RM, Lansing RW, Brown R, Banzett
RB. Reduced tidal volume increases “air hunger” at fixed PCO2 in ventilated quadriplegics. Respir Physiol 1992;90:19-30. https://doi.org/10.1016/0034-5687(92)90131-F
25. Schwartzstein
RM, Manning HL, Woodrow Weiss J, WeinÂberger SE. Dyspnea: A sensory experience
[Internet]. Lung 1990; 168: 185-99. Available from:
http://dx.doi.org/10.1007/bf02719692
26.
Al-Omari A, Alhuqbani WN, Zaidi ARZ, et al. Clinical charÂacteristics
of non-intensive care unit COVID-19 patients in Saudi Arabia: A descriptive
cross-sectional study. J Infect Public Health 2020; 13: 1639-44.
https://doi.org/10.1016/j.jiph.2020.09.003
27.
Guan WJ, Ni ZY, Hu Y, et al. Clinical
Characteristics of Coronavirus Disease 2019 in China. N Engl
J Med 2020;382:1708-20.
https://doi.org/10.1056/NEJMoa2002032
28. Li YC, Bai
WZ, Hashikawa T. The neuroinvasive
potential of SARS-CoV2 may play a role in the respiratory failure of COVID-19
patients. J Med Virol 2020;92:552-5.
https://doi.org/10.1002/jmv.25728
29. Chen T, Wu D, Chen H, et al.
Clinical characteristics of 113 deceased patients with coronavirus disease
2019: retrospective study. BMJ 2020;368:m1091.
https://doi.org/10.1136/bmj.m1091
30. Kaeuffer
C, Le Hyaric C, Fabacher T,
et al. Clinical charÂacteristics and risk factors associated with severe COÂVID-19:
prospective analysis of 1,045 hospitalised cases in
North-Eastern France, March 2020. Euro Surveill
[Internet] 2020;25(48).
http://dx.doi.org/10.2807/1560-7917.ES.2020.25.48.2000895
31. Bhatraju PK, Ghassemieh BJ,
Nichols M, et al. Covid-19 in Critically Ill Patients in the Seattle Region -
Case Series. N Engl J Med. 2020;382:2012-22.
http://dx.doi.org/10.1056/NEJMoa2004500.
32.
Viola L, Russo E, Benni M,
et al. Lung mechanics in type L CoVID-19 pneumonia: a
pseudo-normal ARDS. Transl Med Commun 2020;5:27.
https://doi.org/10.1186/s41231-020-00076-9

| GalerĂa de imágenes | ||
| Mujer joven con afectaciĂłn pulmonar bilateral y alteraciĂłn de la conciencia | ||
Autores: Churin Lisandro |
 |
|